2.4. Szonolumineszcencia
A következő nagyon izgalmas, még a kavitációval összefüggő,
hazánkban is egyre több kutatót foglalkoztató akusztikai jelenség a
szonolumineszcencia. A szonolumineszcencia jelensége fénykibocsátó üreg, vagy
buborék jelenlétét jelenti az ultrahangtérben (22. ábra). Természetesen
általában több ilyen üreg vagy buborék egyidejű jelenlétéről van szó a
szonolumineszcencia említésekor és éppen a monobuborékos stabil rendszerek
kialakítása igényli a legkomolyabb tervező tevékenységet. Látható a 22. ábrán,
hogy a mechanikai erősítő oszlop alatt milyen intenzív kemolumineszcencia
valósul meg ultrahang hatására luminollal. Azonban természetes módon bárki
megtapasztalhatja tiszta csapvíz esetében is ezt a jelenséget egy sötétített
laboratóriumban az ultrahang hatására, megfelelően magas akusztikai
nyomásamplitúdók mellett.
Több próbálkozás történt tehát az egybuborékos rendszerek
életre hívására, azonban a fizikai alapot tekintve mindig állóhullámbeli
levitációt alkalmaznak. Két módszer terjedt el széles körben. Az egyiknél
gömblombikban, álló gömbhullámokkal, a másiknál pedig egy álló hengerben, az
előzetesen vázolt folyadékbeli 6. és 8. ábra szerinti levitátor elrendezésben
történik a buborékok csapdázása a folyadékban, az állóhullám sebességi
csomósíkjaiban, mivel a buborék könnyebb, mint a vivőközeg.
 (A)
(A) (B)
(B) (C)
(C)
22. Ábra: A
szonolumineszcencia megnyilvánulásai (A, kemilumineszcencia, B,
szonolumineszcencia-berendezés, C, szonolumineszcencia levitáló stabil
kavitációs buborék esetén, B berendezésben)
Magát a szonolumineszcencia jelenséget 1934 óta ismeri a
tudományos világ, azonban létezésének igazi tudatosodása egy évtizeddel
ezelőttre tehető. Ekkor dolgozták ki a fent említett gömblombikos és álló hengeres
technikát, amely segítségével a fénykibocsátó üreg huzamos időn keresztül egy
helyben csapdázható a térben (22.B. ábra).
A fénykibocsátás oka a korábban született, viszont a mai
napig konvencionális magyarázat szerint az, hogy több ezer bar és 10-35000K
hőmérséklet alakul ki a kavitációs buborékokban, ami miatt az üregben lévő
atomok gerjesztődése okán foton kibocsátás történik. Ma az egyre gyorsabb
filmfelvételi lehetőségek megjelenésével, egyre többen lézerelmélettel
magyarázzák a jelenséget, mert kiderült, hogy a fénykibocsátás 10
pikoszekundumos sorozatokból tevődik össze. Az utóbbi években egyes mértékadó
tudósok, a még feltáratlan anomáliák miatt, logikailag azt állítják, hogy az
eddigi hőmérséklet és nyomásértékek alábecsültek és bizonyos új elméletek
szerint a fénylő buborékban 15millió°C is kialakulhat, ami a földi napmodell
lehet, melynek során magreakció által, a hélium hidrogénekre bomlik. Persze egy
ilyen arra érdemes kijelentés után több, eltérő impaktfaktú cáfolat között
mazsolázhatunk, de ez az invizíciónál is így volt, csak valamennyire
határozottabban. Végső soron 10-35000K mellett is igen speciális kísérleteket
végezhetünk, hosszútávon stabil akusztikai rendszerrel, ez pedig komoly
fegyvertény.
2.5. Az ultrahang hőhatása, hipertermia
A következőkben tekintsük át az ultrahang minden
felhasználó által ismert hőhatását! A hanghullámok minden anyagban
frekvenciájuktól, a hullám típusától, a hőmérséklettől, illetve az anyag
tulajdonságaitól függő mértékben adszorbeálódnak, a rezgési energia
irreverzibilis hővé alakulása következtében.
Az ultrahang egyik legfantasztikusabb, hozzávetőleg két
évtizedes aktív múltra visszatekintő megrendítő erejű kutatási témájának a
hipertermiának az alapját ez a tény képezi. Az ultrahang hőhatása természetesen
könnyen elegendő a biológiai struktúrák és a kémiai folyamatok
befolyásolásához. Az adszorpció miatt az ultrahang intenzitása a távolsággal
exponenciálisan csökken és az intenzitást a hangtér egy adott pontján, az
alábbi [5]. egyenlettel fejezzük ki:
I = I0 * e –2αx [5]
ahol
(I0) [W/cm2] vagy [dB] a kiindulási, vagy kibocsátott
intenzitás, (I) az aktuális intenzitás [W/cm2] vagy [dB], (α)
az abszorpciós koefficiens [Np/cm=8,7dB/cm], (x) pedig az adott irányban
megtett távolság [cm]. A hőképződés (qv) [K/s] pedig a [6]. egyenlet
szerint:
qv = 2αI/C*m [6]
ahol
(C) a fajhő [kJ/gK], (m) a kezelt anyag tömege [g].
2.5.1. Az abszorpciós koefficiens
Az abszorpció, vagy elnyelődés mindig jellemez egy adott közeget,
környezetet, struktúrát, ami meghatározza a terjedés para
Magától értetődő módon az egyes anyagoknak, így az egyes
szöveteknek is különbözik az abszorpciós koefficiense, amely mellett, ha azt is
figyelembe vesszük, hogy az ultrahang irányítható, koncentrálható, akkor már
érthető, hogy az arra fogékonyak miért jönnek ettől izgalomba. Ugyanis abban az
esetben, ha például a kezelendő szövet előtt nagyobb abszorpciós koefficienssel
rendelkező, vagy érzékenyebb szövetrész található, akkor több kisebb
intenzitású különböző irányokból érkező linearizált, vagy koncentrált hangsugár
alkalmazásával kivitelezhető a kívánt hatású kezelés a megcélzott területen
(23. ábra).
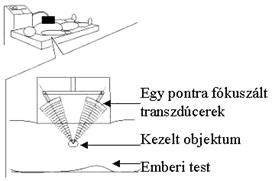
23.
Ábra: Célra irányított ultrahang hipertermia
A 23. ábrán megfigyelhető, hogy a
koncentrált ultrahang sugár segítségével akár az emberi test bármely pontján
elhelyezkedő objektum megcélozható, ahol a hőmérsékletemelkedés hatására a
fehérjék denaturációja következik be. A denaturálódott fehérjék pedig a test
adott zónájából felszívódnak. A mai számítógépes szoftverek segítségével mód
van arra, hogy az egyes hang útjába kerülő objektumok hangsugár törő,
adszorpciós és szóró képességét kikalkulálják, abból a célból, hogy éppen a
megfelelő mennyiségű energia érje el azt a pontot, ahol a kezelendő objektum található.
Természetesen ez a számítógépes sugárútszámításra vonatkozó technika már nem
teljesen „vakrepülés”, a nagy precizitású modellezőszoftverek kifejlesztése
után, de egy újabb, sokkal biztosabb, a pillanatnyi változásokat állandóan
követő és ezek alapján szabályozott, a tanulmányban többször említett
technológia az MR-re (mágneses magrezonanciára) alapozott ultrahangsebészet.
Itt a mágneses magrezonancia képet, mint egy három dimenzióban szkennelt,
állandóan frissített emberi test képet kell elképzelni, amelyen a beavatkozást
végző orvos bejelöli a denaturálandó testterületet és a koncentrált hangsugár
tizedmilli
2.6. Az ultrahang sejtbiológiai hatásai
Nézzük meg először, hogy milyen alapokon nyugszik az
ultrahang sejtbiológiai hatásmechanizmusa! A mai napig, a legfelsőbb alkalmazói
körökben is sok kérdés merül fel a témával kapcsolatban. Ebben a kérdésben,
mivel a szűkebben vett kutatási területemről van szó, megpróbálok korrekt,
szerteágazó és teljes mértékig autentikus választ adni, azonban figyelembe véve
azt, hogy ne haladjam meg a más felhasználási módozatok iránt érdeklődők
türelmi szintjét. Legáltalánosabban Miller et al. (1996) fogalmazták meg az
ultrahang in vitro sejtbiológiai hatásait. Szerintük három alapvető
elemből tevődik össze az ultrahang sejtbiológiai hatásmechanizmusa, melyek a
termikus, a kémiai és a mechanikai mechanizmusok. Morton et al. (1982) szerint
az ultrahangos besugárzás, a szuszpenziókban lévő sejtek líziséhez,
széteséséhez, széttöredezéséhez vezet. Szoros összefüggést mutattak ki a
szuszpendált sejtek roncsolódása és az összegzett kibocsátott alharmonikus
energia közt, amelyről tudjuk, hogy az a stabil kavitációs buborékok
jelenlétének az ismertetőjegye. Azt is kimutatta, hogy amikor elkezdődött az alharmonikus
kibocsátás, akkor kezdtek pusztulni a sejtek, ami megkérdőjelezhetetlen
bizonyíték. A sejtek életerejének felbecsülésére vitális festést alkalmazott
trypon kékkel, hasonlóan a mi általunk alkalmazott metilénkékes vitális festési
eljáráshoz. Harwey et al. (1975) kimutatták, hogy amely sejt közelében
tranziens kavitációs összeomlás történt, ott a sejtek roncsolódása következett
be, melynek kapcsán a sejtfal felrepedt, az endoplazmatikus retikulum kitágult,
a mitokondrium károsodott és más szabálytalan mechanizmusok is felléptek.
Miller et al. (1995) ultrahangsugárzás hatásaként DNS fonal töredezéséről,
Macintosh és Davey (1970) kromoszóma széttöredezésről, Barnett et al. (1988)
kromatída aberrációról, Kaufman (1985) mutagén hatásról, Dooley et al. (1984) a
sejtek makromolekula szintézisének megváltozásáról számolt be. Fu et al. (1980)
vizsgálták a sejtek ultrahangkezelés hatására kialakuló telepképzési
erélyváltozását, amellyel kapcsolatban megállapítják, hogy ez a tulajdonság már
1W/cm2 intenzitású ultrahang besugárzás hatására is megváltozik.
Thacker (1973) a haploid és diploid pékélesztő (Saccharomyces cerevisiae)
sejtek ultrahang besugárzással kapcsolatos túlélését vizsgálta. Tapasztalatai
alapján nem szinkronizált populációk vizsgálatát javasolja, az eltérő fejlődési
stádiumban lévő sejtek eltérő kavitációs érzékenysége miatt. A vizsgálataiból
kapott túlélési görbék, ezáltal nem egy, hanem több fázisúak voltak. Emiatt az
eredményei eltérést mutattak a szokványos exponenciális túlélési görbétől, habár
a kavitációs határon dolgozó kutatók a pusztulási dinamikát az egyszerűség
kedvéért állandó exponenciális lefutásúra veszik. Thacker (1974) egy évvel
később négy genetikai rendszerhez tartozó élesztő sejteket vizsgált, az
ultrahang mutagén hatásának tekintetében. A mitokondriális DNS-ben legtöbbször
mutáció történt a kavitáció hatására. A mutagén hatás gyakorisága pedig
növekedett a hőmérséklet emelkedésével.
2.6.1. A beugárzás hatására keletkező
szonokemikáliák sejtbiológiai hatásai
Fontos megjegyezni, hogy nem csak a szonokémia,
hanem az ultrahang sejtbiológiai hatásainak jelentős része is függ a besugárzás
hatására kialakuló szonokemikáliáktól, melynek legáltalánosabb megnyilvánulási
formája az ultrahang sugárzás hatására kialakuló hidrogén-peroxid (H2O2)és
a (H++OH-) ionok, vagyis a vízbontás folyománya. Hughes
(1961) élesztő sejteket tárt fel akusztikai kavitáció segítségével és arra a
megállapításra jutott, hogy a kavitáció során szabadgyökök keletkeznek, és azok
hozzájárulnak a kavitáció mechanikai roncsoló hatása által kiváltott
sejtpusztuláshoz. Miller et al. (1991) és Riesz és Kondo (1992) szerint a
tranziens kavitáció folyamán képződő szabad gyökök, valamint egyéb szonokémiai
termékek, így például a szonokémiai hidrogén-peroxid jól detektálható, in
vitro ultrahang besugárzást követően, ami hozzájárul a sejtek DNS
károsodásához is. A polimereket (DNS) összekötő hidrogén hidakkal reakcióba lép
a vízbontás hatására kialakult szabadgyök, az (OH-) ion, emiatt az
ultrahang mutagén hatása kerül előtérbe. Miller és Thomas (1994) szintén
kimutatták, hogy a hidrogén-peroxid és egyéb szonokemikáliák, megfelelő
koncentrációban történő termelődése biokémiai változásokat eredményeznek az élő
sejtekben, illetve ehhez a hatáshoz adódik még hozzá a kavitáció által okozott
direkt mechanikai sejtkárosító hatás is. Prise et al. (1989) szerint a
sejtpusztuláshoz szükséges hidrogén-peroxid koncentráció 1mM. Miller és Thomas
(1993) hidrogén-peroxid termelődését és hemolízis megindulását írták le
ugyanazzal a küszöbbel, ahogy a tranziens kavitáció megindult. A kavitáció
következtében a besugárzás 1. perceiben, közel 100%-os sejt lízis (feloldódás)
következett be, viszont a hidrogén-peroxid koncentráció a besugárzás 30.
percében mindössze 10mM volt, ami az előzőek alapján szintén a mechanikai
hatás dominanciáját húzza alá és nem a képződött hidrogén-peroxid pusztító
erejét.
2.6.3. Specifikus sejtbiológiai hatások
Nem szakemberek számára most biztosan elborzasztó
tudományos eredmények ismertetése következik. Azonban felhívom a figyelmet,
hogy a specifikus sejtbiológiai hatások jelen tárgyalásánál, elsősorban az
aktív ultrahang által kiváltott folyamatokra mutatok rá. Ezt jelen esetben
azzal a hasonlattal tudnám megvilágítani, hogy amíg a passzív ultrahang olyan,
mint egy lenge szélfuvallat, addig az aktív olyan, mint egy tomboló orkán. Kim
et al. (1971) és Schnitzler (1973) kromatídák széttöredezéséről
számoltak be, a stabil kavitáció következtében sejtszuszpenziókban. A töredezés
a mitózisos osztódás anafázisban volt a legerőteljesebb, illetve a centromérák
erőteljes töredezésére is felhívták a figyelmet. Azt állítják, hogy az
oszcilláló stabil kavitációs buborékok által kiváltott mikroáramlások okozzák a
kromoszóma aberrációt az ultrahanggal kezelt sejteknél. Hughes és Nyborg (1962)
vizsgálták az Escherichia coli baktériumok ultrahang általi pusztulását,
és ők is azt tapasztalták, hogy nem csak a tranziens, hanem a sokat vitatott
hatású stabil kavitáció esetében is megtörtént a sejtek pusztulása, így ez
alapján ők is azt állítják, hogy a tranziens, összeomló típusú kavitáció nem
elengedhetetlen a sejtek széttöredezéséhez. Kaufman et al. (1977)
bebizonyította, hogy a szuszpendált sejtek ultrahangos besugárzása a sejtek
líziséhez, vagyis széteséséhez, feloldódásához, illetve a sejtek teljes
széttöredezéséhez vezet, aminek fő kiváltója a kavitáció. A lízis közvetlen,
azonnal jelentkező következménye az ultrahangsugárzásnak, nem egy később
expresszálódó hatás. ter Haar et al. (1980) és Li et al. (1977a)
szerint, az ultrahangnak alávetett sejtek esetében, amelyeknél
hőmérsékletnövekedés következik be, és amely sejtek nem pusztulnak el a
mechanikai sérüléseik miatt rögtön, szaporodóképesség vesztés léphet fel.
2.6.4. Az ultrahang biológiai membránokra
gyakorolt hatásai
Nem kell hangsúlyozni a membrántranszport folyamatok
fontosságát. Több évtizede külön membrántranszport szekciója van a Magyar
Biofizikai Társaságnak, önálló tudományterület, különálló konferenciákkal,
folyóiratokkal, tudományos eredményekkel, intézetekkel. Emiatt természetesen a
teljesség igénye nélkül, mindössze érintőlegesen próbálom felhívni a figyelmet,
az ultrahang transzportfolyamatokat befolyásoló hatására. Az ultrahang akkor
befolyásolja a membrántranszport folyamatokat, ha nem a legdrasztikusabb sejtroncsoló
hatásról, hanem a besugárzást túlélt, elszenvedett sejtekről beszélünk. Dinno
et al. (1989) szerint az ultrahang besugárzás megváltoztatja a sejtmembrán
permeabilitását, transzport aktivitását. A sejt elektromos para
2.6.5. Alkalmazott ultrahang fizikai alapjai
Az ultrahang gyakorlati alkalmazására lehet példa,
hogy Rubleson et al. (1975) szerint a mikroorganizmusok ultrahangos
szétroncsolása, a tej pasztőrözésének tekintetében, a konvencionális,
tradicionális sterilizálási és pasztőrözési eljárások kiegészítéseként kerülhet
szóba. A baktériumok kizárólag ultrahangos elpusztítása nehézkes, de az
ultrahang fel tudja erősíteni a konvencionális hőkezelés hatását, amely
folyamat így felgyorsul, mivel a baktérium plakkok a besugárzás hatására
diszpergálódnak, így javul a denaturálandó anyagok felé irányuló hőtranszport.
Ordonez et al. (1984) hőkezeléssel kombinálta az ultrahang besugárzást, és azt
állapította meg, hogy a baktérium sejtek érzékenyebbek a hőkezelésre, ha
ultrahangnak is ki vannak téve. Hurst et al. (1995) javasolja a hőkezelés
ultrahangkezeléssel való szimultán alkalmazás elnevezésének a
termoultraszonikáció kifejezést. Megállapította, hogy a termoultraszonikáció
eredményesebben alkalmazható a sejtek elpusztítására, mint a hő vagy az
ultrahang kezelés önálló alkalmazása.
2.6.6. Az ultrahang sejtbiológiai hatásait
befolyásoló fizikai tényezők
Az ultrahang sejtbiológiai hatásainak fizikai
befolyásoló tényezői közül a legfontosabbakat az alábbiakban próbáltam
rendszerezni. Számos fizikai, kémiai és biológiai befolyásoló tényező hat a
hangtérben, amelyeknek eredője a sejtbiológiai hatás. Fontos e helyen leszögezni
azt, hogy amennyiben a hangtér fizikai para
2.6.7. Az ultrahangsugárzás hatásának
bemutatása
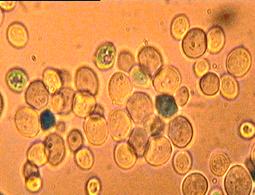
A 24. ábra egy in vitro ultrahangbesugárzás
hatását mutatja Saccharomyces cerevisiae élesztőgombára.
 (A)
(A) 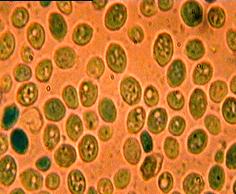 (B)
(B)
24. Ábra: Élesztőgomba vitalitás változása (A. Kiindulási állapot,
B. 210 másodperces kezelés utáni állapot (Forrás: Lőrincz, A., 2002))
Az ábrákon metilénkék vitális festés segítségével
végzett élősejtszám meghatározás eredménye látható. A kezeléseket 1117kHz
(1,117MHz) frekvencián 9W/cm2 teljesítmény mellett 20°C-on végeztük
20ml 5,37*107/ml szuszpenzió koncentráció mellett a 25. ábrán
látható ultrahangrendszer és a 26. ábrán látható kezelőedény segítségével. A
rendszert teljes egészében mi terveztük és készítettük. Segítségével mérhető a
hangsebesség, az abszorpciós koefficiens folyamatosan, akár átfolyó rendszerben
is, illetve a passzív mérésekkel párhuzamosan aktív ultrahang besugárzás is
történhet. Így folyamatosan elemezhetővé válik az aktív ultrahang hatása a
kezelt, például biológiai anyagokra. A műszerrel egyedülállóan vizsgálhatóak az
egyes akusztikai jelenségek fizikai és biofizikai hatásai.

25. Ábra: Az ultrahang
rendszer (vezérlő és adatfeldolgozó computer, ultrahang erősítő,
frekvenciagenerátor, kezelőedények (Forrás: Lőrincz, A.,
2003))
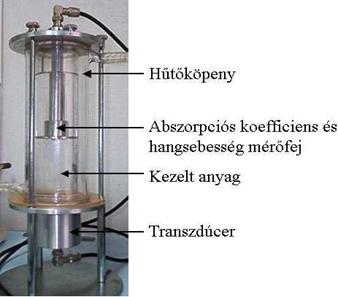
26.
Ábra: Ultrahang kezelőedény (Forrás:
Lőrincz, A., 2003)
A metilénkékkel végzett vitális festés esetén a kék
sejt pusztultnak tekinthető, mivel dehidrogenázokkal való reakciója folytán a
festék az élő sejtekben színtelenné válik. Látható, hogy a 210. percben az
adott mikroszkóp látótér alatti sejtek 100%-a pusztultnak vehető, illetve nagy
részükön világos foltok találhatóak, melyek a leszakadt sarjadzó sejtek,
illetve a sejtfal kiszakadások nyomai. A további kezelés az élesztőgomba
sejtekből protoplazmacseppeket, majd sejttörmeléket alakít ki.